Генеалогический метод используется для. Генеалогический метод исследования наследственности человека - контрольная работа. Определение генотипа пробанда
К методам, широко используемым при изучении генетики человека, относятся генеалогический, популяционно-статистический, близнецовый, метод дерматоглифики, цитогенетический, биохимический, методы генетики соматических клеток.
Генеалогический метод
В основе этого метода лежит составление и анализ родословных. Этот метод широко применяют с древних времен и до наших дней в коневодстве, селекции ценных линий крупного рогатого скота и свиней, при получении чистопородных собак, а также при выведении новых пород пушных животных. Родословные человека составлялись на протяжении многих столетий в отношении царствующих семейств в Европе и Азии.
Как метод изучения генетики человека генеалогический метод стали применять только с начала XX столетия, когда выяснилось, что анализ родословных, в которых прослеживается передача из поколения в поколение какого-то признака (заболевания), может заменить собой фактически неприменимый в отношении человека гибридологический метод.
При составлении родословных исходным является человек - пробанд, родословную которого изучают. Обычно это или больной, или носитель определенного признака, наследование которого необходимо изучить. При составлении родословных таблиц используют условные обозначения, предложенные Г. Юстом в 1931 г. (рис. 7.24). Поколения обозначают римскими цифрами, индивидов в данном поколении - арабскими.
С помощью генеалогического метода может быть установлена наследственная обусловленность изучаемого признака, а также тип
Рис. 7.24. Условные обозначения при составлении родословных (по Г. Юсту) его наследования (аутосомно-доминантный, аутосомно-рецессив- ный, Х-сцепленный доминантный или рецессивный, Y-сцепленный). При анализе родословных по нескольким признакам может быть выявлен сцепленный характер их наследования, что используют при составлении хромосомных карт. Этот метод позволяет изучать интенсивность мутационного процесса, оценить экспрессивность и пене- трантность аллеля. Он широко используется в медико-генетическом консультировании для прогнозирования потомства. Однако необходимо отметить, что генеалогический анализ существенно осложняется при малодетности семей.
Для аутосомного типа наследования в целом характерна равная вероятность встречаемости данного признака как у мужчин, так и у женщин. Это обусловлено одинаковой двойной дозой генов, расположенных в аутосомах у всех представителей вида и получаемых от обоих родителей, и зависимостью развивающегося признака от характера взаимодействия аллельных генов.
При доминировании признака в потомстве родительской пары, где хотя бы один родитель является его носителем, он проявляется с большей или меньшей вероятностью в зависимости от генетической конституции родителей (рис. 7.25).
Если анализируется признак, не влияющий на жизнеспособность организма, то носители доминантного признака могут быть как гомо-, так и гетерозиготами. В случае доминантного наследования какого-то патологического признака (заболевания) гомозиготы, как правило, нежизнеспособны, а носители этого признака - гетерозиготы.
Таким образом, при аутосомно-доминантном наследовании признак может встречаться в равной мере у мужчин и у женщин и прослеживается при достаточном по численности потомстве в каждом поколении по вертикали. Анализируя родословные, необходимо помнить о возможности неполного пенетрирования доминантного аллеля, обусловленной взаимодействием генов или факторами среды. Показатель пенетрантности может быть вычислен как отношение фактического числа носителей признака к числу ожидаемых носителей этого признака в данной семье. Необходимо также помнить, что некоторые заболевания проявляются не сразу с момента рождения

Рис. 7.25. Вероятность появления потомков с доминантным признаком от различных супружеских пар (/ - III)
ребенка. Многие болезни, наследуемые по доминантному типу, развиваются лишь в определенном возрасте. Так, хорея Гентингтона клинически проявляется к 35-40 годам, поздно проявляется и поли- кистоз почек. Поэтому при прогнозировании подобных заболеваний в расчет не принимаются братья и сестры, не достигшие критического возраста.
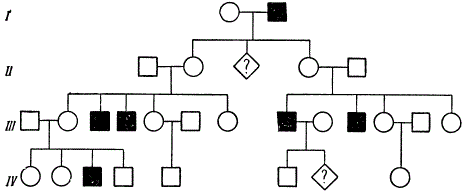
Первое описание родословной с аутосомно-доминантным типом наследования аномалии у человека было дано в 1905 г. В ней прослеживается передача в ряду поколений брахидактилии (короткопалое™). На рис. 7.26 приведена родословная с этой аномалией. На рис. 7.27 изображена родословная с ретинобластомой в случае неполной пенетрантности.
Рецессивные признаки проявляются фенотипически лишь у гомозигот по рецессивным аллелям. Эти признаки, как правило,

Рис. 7.26. Родословная (X) при аутосомно-доминантном типе наследования (бра- хидактилия - Б)

Рис. 7.27.
Рис. 7.28. Вероятность появления потомков с рецессивным признаком от различных супружеских пар
обнаруживаются у потомков фенотипически нормальных родителей - носителей рецессивных аллелей. Вероятность появления рецессивного потомства в этом случае равна 25%. Если один из родителей имеет рецессивный признак, то вероятность проявления его в потомстве будет зависеть от генотипа другого родителя. У рецессивных родителей все потомство унаследует соответствующий рецессивный признак (рис. 7.28).
Для родословных при аутосомно-рецессивном топе наследования характерно, что признак проявляется далеко не в каждом поколении. Чаще всего рецессивное потомство появляется у родителей с доминантным признаком, причем вероятность появления такого потомства возрастает в близкородственных браках, где оба родителя могут являться носителями одного и того же рецессивного аллеля, полученного от общего предка. Примером аутосомно-рецессивного наследования является родословная семьи с псевдогипертрофиче- ской прогрессивной миопатией, в которой часты близкородственные браки (рис. 7.29). Обращает внимание распространение заболевания в последнем поколении по горизонтали.
Гены, расположенные в Х-хромосоме и не имеющие

Рис. 7.29. Родословная при аутосомно-рецессивном типе наследования (псевдоги- пертрофическая прогрессирующая миопатия) аллелей в Y-хромосоме, представлены в генотипах мужчин и женщин в разных дозах. Женщина получает две свои Х-хромосомы и соответствующие гены как от отца, так и от матери, а мужчина наследует свою единственную Х-хромосому только от матери. Развитие соответствующего признака у мужчин определяется единственным аллелем, присутствующим в его генотипе, а у женщин он является результатом взаимодействия двух аллельных генов. В связи с этим признаки, наследуемые по Х-сцепленному типу, встречаются в популяции с разной вероятностью у мужского и женского пола.
При доминантном Х-сцепленном наследовании признак чаще встречается у женщин в связи с большей возможностью получения ими соответствующего аллеля либо от отца, либо от матери. Мужчины могут наследовать этот признак только от матери. Женщины с доминантным признаком передают его в равной степени дочерям и сыновьям, а мужчины - только дочерям. Сыновья никогда не наследуют от отцов доминантного Х-сцепленного признака.
Примером такого типа наследования служит описанная в 1925 г. родословная с фолликулярным кератозом - кожным заболеванием, сопровождающимся потерей ресниц, бровей, волос на голове (рис. 7.30). Характерным является более тяжелое течение заболевания у гемизиготных мужчин, чем у женщин, которые чаще всего являются гетерозиготами.
При некоторых заболеваниях наблюдается гибель мужчин-геми- зигот на ранних стадиях онтогенеза. Тогда в родословных среди пораженных должны быть только женщины, в потомстве которых отношение пораженных дочерей, здоровых дочерей и здоровых сыновей равно 1:1:1. Мужские доминантные гемизиготы, не погибающие на очень ранних стадиях развития, обнаруживаются в самопроизвольных абортах или среди мертворожденных. Такими особенностями наследования у человека характеризуется пигментный дерматоз.
Характерной особенностью родословных при данном типе наследования является преимущественное проявление признака у гемизиготных мужчин, которые наследуют его от матерей

Рис. 7.30. Родословная при Х-сцепленном доминантном типе наследования (фолликулярный кератоз)

Рис. 7.31. Родословная при Х-сцепленном рецессивном типе наследования (гемофилия типа А)
с доминантным фенотипом, являющихся носительницами рецессивного аллеля. Как правило, признак наследуется мужчинами через поколение от деда по материнской линии к внуку. У женщин он проявляется лишь в гомозиготном состоянии, вероятность чего возрастает при близкородственных браках.
Наиболее известным примером рецессивного Х-сцепленного наследования является гемофилия. Наследование гемофилии типа А представлено в родословной потомков английской королевы Виктории (рис. 7.31).
Другим примером наследования по данному типу является дальтонизм - определенная форма нарушения цветоощущения.
Наличие Y-хромосомы только у представителей мужского пола объясняет особенности Y-сцепленного, или голандрического, наследования признака, который обнаруживается лишь у мужчин и передается по мужской линии из поколения в поколение от отца к сыну.

Рис. 7.32. Родословная при Y-сцепленном (голандрическом) типе наследования
Одним из признаков, Y-сцепленное наследование которого у человека все еще обсуждается, является гипертрихоз ушной раковины, или наличие волос на внешнем крае ушной раковины. Предполагают, что в коротком плече Y-хромосомы кроме этого гена находятся гены, определяющие мужской пол. В 1955 г. у мыши описан определяемый Y-хромосомой трансплантационный антиген, названный HY.
Возможно, он является одним из факторов половой дифференцировки мужских гонад, клетки которых имеют рецепторы, связывающие этот антиген. Связанный с рецептором антиген активизирует развитие гонады по мужскому типу (см. разд. 3.6.5.2; 7.1.2).
Этот антиген в процессе эволюции остался почти неизменным и встречается в организме многих видов животных, в том числе
и человека. Таким образом, наследование способности к развитию гонад по мужскому типу определяется голандрическим геном, расположенным в Y-хромосоме (рис. 7.32).
У человека как объекта генетического исследования почти нет никаких преимуществ перед другими объектами.
Напротив, много препятствий, затрудняющих изучение его генетики: 1) невозможность произвольного скрещивания в эксперименте; 2) позднее наступление половой зрелости; 3) малое число потомков в каждой семье; 4) невозможность уравнивать условия жизни для потомства; 5) отсутствие точной регистрации проявления наследственных свойств в семьях и отсутствие гомозиготных линий; 6) большое число хромосом; 7) и самым главным затруднением изучения генетики человека в капиталистическом обществе является социальное неравенство, что затрудняет реализацию наследственных потенций человека.
Несмотря на указанные затруднения, генетика разработала некоторые методы, которые позволяют шаг за шагом изучать наследственность и наследование у человека. Существует несколько методов исследования: генеалогический, цитогенетический, близнецовый, онтогенетический и популяционный.
Следует иметь в виду, что любой признак, независимо от того, является ли он признаком дикого типа, т. е. относится к норме, или связан с каким-либо заболеванием, может служить моделью для изучения наследственности. Оградить человека от наследственных болезней или поражения его наследственности так же важно, как и выяснить наследование нормы. В настоящее время генетические методы разработаны главным образом в отношении морфологических признаков, которые генетически определяются достаточно четко (брахидактилия, альбинизм, дальтонизм, пятнистость кожи и волос и т. д.).
Генетическое исследование психических свойств все еще остается проблематичным, так как для них не найдены элементарные критерии признака в генетическом смысле. Почти все признаки Психической и творческой деятельности человека настолько комплексны и сложны, а также в сильной степени обусловлены внешними, в том числе и социальными, факторами, что генетический анализ этих свойств пока трудно осуществим, хотя наследственная их обусловленность не вызывает сомнения.
Можно сказать, что значительное большинство признаков, характеризующих вид Homo sapiens, может изучаться как количественные и сложные физиологические признаки, т. е. признаки, не проявляющие дискретного характера в онтогенезе. Эти признаки контролируются системой генотипа (полигенно). И пока эта система не разгадана хотя бы на примере просто организованных организмов, проблема признаков поведения остается малодоступной для генетического анализа. Напротив, мутантные признаки, выходящие за границы характеристики видовых признаков, служат хорошими генетическими моделями изучения наследственности и наследования в норме.
На дискретные мутантные признаки нельзя смотреть как на признаки только патологические, якобы не имеющие приспособительного значения. Возможно, что само появление человека с развитыми полушариями коры головного мозга, вертикальным положением тела, дискретной речевой сигнализацией является следствием крупных мутаций. В пользу этого свидетельствует очень
короткий промежуток времени эволюции человека, за который мелкие мутации вряд ли могли накопиться в таком количестве и дать такой значительный эволюционный эффект. Разумный человек для природы столь же «необычен», как домашняя курица, несущая 365 яиц в год вместо 10-15, или рекордистка-корова, дающая 16 тыс. кг молока в год вместо 600-700 кг.
Разделение признаков на нормальные и мутантные применительно к человеку и животным необходимо для познания эволюции человека и патологических явлений.
Совокупность видовых признаков человека и животных определяется системой генотипа, сложившейся под влиянием всех факторов отбора в процессе эволюции. Мутации, пребывающие в гетерозиготном состоянии у человека, по-видимому, так же необходимы, как и у животных, для поддержания их в популяции.
Самым опасным в разработке научных методов исследования животных и человека, особенно его способностей, является антропоморфический момент, т. е. выдача желаемого за действительность.
Генеалогический метод
Анализ наследования человека на основе составления родословной - генеалогии был предложен Ф. Гальтоном.
Генеалогический метод представляет собой изучение наследования свойств человека по родословным (педигри). Данный метод применим, если известны прямые родственники - предки обладателя наследственного признака (пробанда) по материнской и отцовской линиям в ряду поколений и имеется достаточное число потомков в каждом поколении, или в том случае, когда имеются данные по достаточному числу разных семей, позволяющему выявить сходство родословных. Данные по совокупности сходных родословных подвергают статистической обработке.
Получившая наибольшее распространение система обозначения родословных человека была предложена Г. Юстом в 1931 г.
На основе большого числа проанализированных семей составляют родословные и производят математические расчеты соответственно типу наследования того или иного признака - доминантному или рецессивному, часто и не часто встречающейся мутации, сцепленному или не сцепленному с полом и т. д. Здесь мы не будем касаться приложения математического метода к данному анализу, отметим только, что весь этот формальный анализ основан на элементарных генетических закономерностях наследования.
Схемы родословной наследования дохминантного аутосомного гена, определяющего какой-либо признак, например заболевание (хондродистрофическая карликовость, буллезный эпидермолиз - свойство кожи образовывать большие пузыри при небольших травмах, ретинобластома и т. д.), или морфологический недостаток, например короткопалость (брахидактилия - отсутствие двух дистальных фаланг в пальцах).

Наследование признаков, определяемых рецессивными генами (рецессивное наследование), анализируется несколько сложнее, при составлении схем родословных.
Например, двоих в семье, появление двоих больных детей равно произведению вероятностей, т. е. 0,25 X 0,25, т. е. 6,25%.

Часто встречающиеся рецессивные аутосомные гены при условии, если носители их (аа) способны вступать в брак и давать потомство, будут находиться в высокой концентрации в популяции. В таком случае становятся очень вероятными браки аа X Аа, в потомстве от которых наследование данного признака будет имитировать наследование по доминантному типу 1:1. Однако, зная тип наследования и проявления тех и других генов даже в случае малочисленных семей, но при достаточном числе таких семей, можно установить истинный характер наследования.
Наследование генов, полностью сцепленных с полом, т. е. находящихся в негомологичных сегментах, и частично сцепленных с полом - локализованных в гомологичных сегментах X- и Y-xpoмосом, подчиняется установленным для половых хромосом закономерностям. Для доминантных и рецессивных генов это наследование будет определяться по-разному в зависимости от того, где данный ген локализован - в гомологичном или негомологичном сегменте X- и Y-хромосомы и каким образом он передается. Так, доминантный ген, вызывающий перепончатость пальцев, находящийся в негомологичном сегменте Y-хромосомы, наследуется от отцов и проявляется только у мужчин.
Для частично сцепленных с полом доминантных генов, находящихся в гомологичных сегментах половых хромосом, анализ несколько более затруднен, но также возможен. Примером сцепленного с полом наследования рецессивного признака является наследование гемофилии. В передаче этого признака в поколениях имеется прерывность; пораженные мужчины являются потомками здоровых матерей, которые были гетерозиготами по данному гену; больные гемофилией женщины могут быть потомками больного отца и больной или здоровой матери.
У человека найдено около 50 сцепленных с полом рецессивных генов. Интересно, что около половины из них обусловливают заболевание глаз. Уже с давних времен было известно, что степень передачи наследственных признаков в родственных (инбридинг) и неродственных браках (аутбридинг) различна. После того, как генетика установила закономерности более частого проявления рецессивных генов при инбридинге, нет необходимости пространно доказывать вред родственных браков. Чем выше коэффициент инбридинга, тем больше вероятность появления наследственных болезней в поколениях. В разных странах среди разных народов и классов общества, а также в разные эпохи родственные браки (между двоюродными, троюродными братьями и сестрами) встречаются с разной частотой. Так, например, в деревнях на островах Фиджи количество родственных браков достигает 29,7%, в некоторых кастах Индии - 12,9, в Японии (Нагасаки) - 5,03, в Голландии - 0,13-0,159, в Португалии- 1,40, в США (Балтимора) - 0,05%, и т. д. Процент родственных браков колеблется в отдельных районах одной и той же страны в зависимости от уклада жизни.
Вредность родственных браков мало заметна в отдельных родословных, но при сравнительном статистическом анализе болезней и смертностей она выступает с полной очевидностью.

Яркий пример выявления рецессивного гена при родственном браке.

В этой родословной родство поддерживается через бракосочетание сибсов (братья - сестры) разной степени родства. От двух родственных браков (четвероюродные сибсы) появилось в одной семье 4 ребенка из 8, а в другой - 2 из 5, страдающих наследственной амавротической идиотией. К. Штерн предполагает, что один из двух общих предков этих линий передал данный рецессивный ген через три поколения каждому из четырех родителей.
Иногда заболевание и смертность детей от родственных браков превышают на 20-30% таковые от неродственных браков. Очевидно, что причина рассматриваемого явления генетическая, а именно: большая вероятность проявления наследственных заболеваний и смертности в результате гомозиготизации рецессивных генов, определяющих физиологические недостаточности и смертность (летальные и полулетальные гены).
Итак, генеалогический метод является весьма ценным методом, однако его значение в исследованиях тем больше, чем точнее и глубже составлены родословные. По мере роста цивилизации и более точной регистрации родословных роль этого метода в генетике человека будет возрастать.
Близнецовый метод
Близнецами называют потомство, состоящее из одновременно родившихся особей у одноплодных животных (человек, лошадь, крупный рогатый скот, овцы и др.).
Близнецы могут быть однояйцевыми и разнояйцевыми.
Идентичные, или однояйцевые, близнецы (ОБ) развиваются из одного яйца, оплодотворенного одним сперматозоидом, когда из зиготы вместо одного зародыша возникают два или более (полиэмбриония). В силу того, что митотическое деление зиготы дает два равнонаследственных бластомера, однояйцевые близнецы, сколько бы их ни развивалось, должны быть наследственно идентичны и одного пола. Это явление представляет собой пример бесполого, а точнее, вегетативного размножения животных.
Разнояйцевые близнецы (РБ) развиваются из одновременно овулировавших разных яйцеклеток, оплодотворенных разными сперматозоидами. И так как разные яйцеклетки и сперматозоиды могут нести различные комбинации генов, то разнояйцевые близнецы могут быть наследственно столь же разными, как и дети одной и той же супружеской пары, родившиеся в разное время. Разнояйцевые близнецы могут быть одного (РБо) или разного пола (РБр).
Чаще в литературе вместо термина «разнояйцевые близнецы» (РБ) употребляют термин «двуяйцевые близнецы» (ДБ), так как двойни встречаются чаще. Однако термин «разнояйцевые близнецы» лучше подчеркивает разницу между ОБ и РБ; однояйцевые близнецы также чаще рождаются двойнями.
Судя по данным, полученным на млекопитающих, для объяснения образования ОБ у человека может быть несколько гипотез:
- расхождение бластомеров при первом дроблении зиготы и раздельное развитие зародыша из этих бластомеров;
- разделение группы клеток на стадии бластоциста (до гаструляции);
- разделение зародышей на ранней стадии гаструляции. Наиболее вероятным путем предполагают второй.

Число близнецов в одних родах у человека колеблется: чаще всего встречаются двойни, реже тройни, еще реже - четверни, совсем редко - пятерни. По данным И. И. Канаева, за последние 150 лет в США установлено четыре случая родов пятерни, в Канаде - два случая. Факт рождения ОБ - пятерни девочек, доживших до взрослого состояния, - известен в семье канадского фермера Дионн (1934 г.). Рассчитано, что пятерни рождаются один раз на 54 700 816 родов, шестерни - на 4712 млн. родов, семерни известны только как исключение. В среднем частота рождения близнецов составляет 1% с колебаниями в пределах 0,5-1,5%. Близнецы менее жизнеспособные, и поэтому их количество при рождении меньше, чем при зачатии, а во взрослом состоянии меньше, чем при рождении.
Расчет частоты ОБ по отношению к РБ делается исходя из теоретического соотношения однополых и разнополых пар РБ при рождении близнецов: 25%♀♀ + 50%♀♂ + 25%♂♂ вычитание числа пар разного пола из общего числа всех пар одинакового пола (мужского и женского) даст разницу, составляющую число пар ОБ, которая в среднем колеблется от 21 до 33,4% всех близнецов.
Для использования близнецов в генетических исследованиях очень важно точно диагностировать тип ОБ и тип РБ. Диагностика производится на основании следующих критериев:
- ОБ обязательно одного пола, РБ могут быть как одного пола, так и разных полов;
- ОБ имеют, как правило, один общий хорион, РБ - разные хорионы;
- реципрокная трансплантация тканей у ОБ столь же успешна, как и автотрансплантация, у РБ она невозможна;
- наличие сходства (конкордантности) у ОБ и несходства (дискордантности) у РБ по многим признакам.
Для диагностики следует выбирать признаки, четко наследующиеся и менее всего подверженные изменению под влиянием факторов среды; к таким признакам относятся группы крови, пигментация глаз, кожи и волос, кожный рельеф (отпечатки кончиков пальцев, ладоней, ступней и др.). Если по одному-двум таким признакам выявлено различие близнецов, то они, как правило, являются РБ.
Все сомнительные случаи диагностики близнецов могут быть вызваны либо нарушением развития одного из партнеров ОБ, либо сходством родителей по ряду признаков. Однако последнее встречается чрезвычайно редко. Следует заметить, что нарушение развития одного из партнеров ОБ обычно объясняют неодинаковым действием факторов внутриутробной жизни и возникновением соматических мутаций на ранних стадиях эмбрионального развития, до закладки органов. Различного рода генные и хромосомные перестройки, моносомия и другие мутации, возникающие у одного из партнеров, способны вызвать значительные различия в фенотипе ОБ. Поэтому необходимо учитывать возможность соматических мутаций у ОБ в раннем эмбриогенезе.
Согласно обобщениям И. И. Канаева, изложенным в его превосходной монографии сущность близнецового метода в генетике сводится к следующим положениям:
1) пара ОБ имеет тождественную комбинацию, пара РБ - разные комбинации генотипов родителей;
2) для обоих партнеров одной пары ОБ внешняя среда может оказаться одинаковой, а для другой - разной. Если партнеры ОБ в течение жизни испытывают разное влияние, то это приведет к внутрипарному различию. Отсюда пары могут быть с внутрипарной одинаковой и внутрипарной разной средой.
Сравнение ОБ с одинаковой средой с ОБ с разной средой открывает возможность судить о роли влияния среды на внутрипарные различия близнецов в течение всей жизни. Сравнение ОБ с одинаковой средой и РБ с одинаковой средой позволяет выяснить роль наследственного фактора. Такого рода изучение проводят на большой выборке и обрабатывают статистически.
Исходя из разности генетического происхождения ОБ и РБ вытекает, что если по одним и тем же признакам нет различия у ОБ и есть таковые у РБ, то очевидно, что данные различия признаков у последних обусловлены наследственными факторами. Если же внутрипарные различия в тех же признаках встречаются у одного и другого типа близнецов, то очевидно, что они могут быть вызваны факторами среды. Из данных дискордантности у ОБ и РБ по ряду морфологических признаков, видно, что внутрипарное различие у РБ встречается во много раз чаще, чем у ОБ.

Представлены некоторые данные С. Рида относительно сравнительной частоты патологии у второго партнера в случае заболевания одного из близнецов.

В процентах показана частота конкордантности заболеваний у двух типов близнецов, из него видно, что если один партнер заболел одной из указанных болезней, то вероятность заболевания второго у ОБ значительно выше, чем у РБ. В. П. Эфроимсон, анализируя данные по частоте контордантных пар, совершенно правильно указывает, что высокая Наследственная предрасположенность ОБ к заболеваниям проявляется при наличии провоцирующего фактора; без него этот процент будет значительно ниже.
Близнецовый метод дает возможность с наибольшей точностью выяснить наследственную предрасположенность человека к ряду заболеваний и свойств. Другими методами очень трудно или почти невозможно исследовать многие инфекционные и опухолевые заболевания, воспаления кожи и различных органов, а также характеристики нормальной нервной деятельности человека.
При использовании близнецового метода приходится учитывать условия совместного и раздельного воспитания в жизни партнеров, социальные условия, в которых они находятся, и т. д. Тем не менее близнецовый метод позволяет наиболее точно определить, коэффициент наследуемости разных признаков, а также судить о гетерогенности популяции по изучаемым генам и вычленять роль среды в определении изменчивости изучаемых признаков.
Цитогенетический метод
Цитогенетическим методом в генетике человека обычно называют цитологический анализ кариотипа человека в норме и патологии.
Правильнее этот метод называть цитологическим, а не цитогенетическим, поскольку генетический анализ путем скрещивания у человека исключен, и носители хромосомных нарушений если выживают, то оказываются, как правило, бесплодными. Однако изредка в отношении некоторых хромосомных нарушений удается сочетать цитологический метод с генеалогическим и устанавливать связь фенотипического эффекта с определенным типом хромосомных изменений. В силу этих обстоятельств можно сохранить принятый в литературе термин «цитогенетический метод» в изучении генетики человека. В тех же случаях, где такого параллелизма исследовании не ведется, применение данного термина неправомочно.
Цитогенетическим методом исследуют различного рода гетероплоидию и хромосомные перестройки в соматических тканях человека, вызывающие различные фенотипические отклонения от нормы.
Чаще всего этот метод применяют на культуре ткани. Он позволяет учитывать крупные аномалии хромосом, возникающие как в половых, так и соматических клетках. Оказалось, что у человека, так же как и у животных, довольно часто возникают трисомики и моносомики по различным парам хромосом вследствие нерасхождения аутосом и половых хромосом в мейозе. Трисомия и моносомия по половым хромосомам у человека обнаруживаются на основе анализа полового хроматина.
В ходе относительно продолжительного индивидуального развития человека в клетках различных тканей накапливаются аномалии хромосом (хромосомные перестройки, а также изменение числа хромосом). Ткани организма представляют собой разнообразные популяции генетически различающихся клеток, в которых с возрастом концентрация клеток с патологическими ядрами возрастает. В этом случае цитогенетический метод позволяет изучать старение тканей на основе исследования структур клеток в возрастной динамике «популяции» соматических и генеративных тканей.
Поскольку частота возникновения хромосомных аномалий зависит от влияния на организм разнообразных мутагенов (ионизации, химических агентов - фармакологических препаратов, газового состава среды и др.), то цитогенетический метод позволяет устанавливать мутагенное действие факторов внешней среды на человека.
Применение цитогенетического метода особенно расширилось в связи с открытием причин ряда физических и психических заболеваний - так называемых хромосомных болезней.
Существует несколько заболеваний человека, например болезнь Клайнфельтера, Шерешевского-Тернера, Дауна и др., причины которых долго оставались неизвестными, пока цитологическими методами у таких больных не были обнаружены хромосомные аномалии.
Больные мужчины с синдромом Клайнфельтера характеризуются недоразвитием гонад, дегенерацией семенных канальцев, умственной отсталостью, непропорциональным ростом конечностей и т. д. У женщин встречается синдром Шерешевского-Тернера. Он проявляется в замедлении полового созревания, недоразвитии гонад, отсутствии менструаций, бесплодии, малом росте и в других Патологических признаках.
Оказалось, что оба эти синдрома у потомков являются следствием нерасхождения половых хромосом при образовании гамет родителей. Вследствие нерасхождения Х-хромосом у женского гомогаметного) пола в процессе мейоза могут возникать гаметы двумя Х-хромосомами, т. е. XX + 22 аутосомы, и без Х-хромосом, т. е. 0 + 22; у мужского (гетерогаметного) пола соответственно гаметы XY + 22 и 0 + 22. В случае оплодотворения таких яйцеклеток нормальными сперматозоидами (X + 22 или Y + 22) возможно образование следующих классов зигот: XXX + 44, 0Х + 44 и XXY + 44, 0Y + 44.
Из этого следует, что число хромосом у зигот разного происхождения может колебаться от 47 до 45, причем особи 0Y + 44, очевидно, не выживают, так как ни разу не были найдены. Хромосомный набор XXY + 44 присущ мужчине с синдромом Клайнфельтера (мужская интерсексуальность), хромосомные наборы Х0 + 44 и XXX + 44 имеют женщины с синдромом Шерешевского-Тернера.
При дальнейшем анализе больных с разными синдромами выяснилось, что вследствие нерасхождения половых хромосом могут возникать разного типа хромосомные аномалии, в частности полисомия. Встречаются, например, мужчины с такими наборами хромосом: XX Y, XXX Y, ХХХХ Y, а женщины - XXX, ХХХХ.
Особенность роли половых хромосом в детерминации пола у человека в случае их нерасхождения, в отличие от дрозофилы, проявилась в том, что набор хромосом XX Y всегда определяет мужской пол, а набор Х0 - женский. При этом увеличение числа Х-хромосом в сочетании с одной Y-хромосомой не изменяет определение мужского пола, а лишь усиливает синдром Клайнфельтера. Трисомия, или полисомия по Х-хромосоме, у женщин также часто вызывает заболевания, сходные с синдромом Шерешевского-Тернера.
Заболевания, вызванные нарушением нормального числа половых хромосом, диагностируются цитологическим методом - анализом полового хроматина. В тех случаях, когда в тканях мужчин имеется нормальный набор половых хромосом - XY, половой хроматин в клетках не обнаруживается. У нормальных женщин - XX - он обнаруживается в виде одного тельца. При полисомии по Х-хромосомам у женщин и мужчин количество телец полового хроматина всегда на единицу меньше числа Х-хромосом, т. е. n x = n·Х - 1. Так, в клетках мужчин с синдромом Клайнфельтера при наборе XX Y имеется одно тельце полового хроматина, при наборе XXXY - два, при наборе XXXXY - три; у женщин с синдромом Шерешевского-Тернера соответственно: Х0 - нет тельца, XXX - два тельца, ХХХХ - три тельца полового хроматина, и т. д. Предполагается, что в каждой такой зиготе генетически активна лишь одна из Х-хромосом. Остальные хромосомы переходят в гетеропикнотическое состояние в виде полового хроматина.
Причины этой закономерности не выяснены, однако предполагается, что она связана с нивелированием действия генов половых хромосом у гетеро- и гомогаметного пола.
Как мы знаем, нерасхождение хромосом может происходить не только в мейозе, но и в соматических клетках в течение всего эмбриогенеза животного начиная с первых дроблений яйца. В силу последнего среди людей при нарушении расхождения половых хромосом могут появиться больные мозаики-женщины и мозаики-мужчины. Так, например, описаны мозаики следующих типов: двойные: Х0/XX, Х0/XXX и X0/XY, X0/XYY, тройные: Х0/ХХ/ХХХ, XX/X0/XY, а также четверные мозаики, когда соматические клетки одного человека содержат четыре разных набора хромосом.
Кроме рассмотренного типа болезней, вызванных изменением числа половых хромосом в зиготе, хромосомные болезни могут быть вызваны нерасхождением аутосом и разного рода хромосомными перестройками (транслокациями, делециями). Так, например, у детей с врожденной идиотией - болезнью Дауна, сопровождающейся малым ростом, широким круглым лицом, близко расположенным узкими глазными щелями и полуоткрытым ртом, была обнаружена трисомия по 21 хромосоме. Установлено, что частота встречаемости болезни Дауна у новорожденных зависит от возраста матерей.
С врожденными хромосомными аномалиями связывают весьма разнообразные болезни. Поэтому цитогенетический метод приобретает важное значение в этиологии болезней человека.
Популяционный метод
Популяционный метод позволяет изучать распространение отдельных генов или хромосомных аномалий в человеческих популяциях.
Популяционный метод основывается на математических методах. Для анализа генетической структуры популяции необходимо обследовать большую по размеру выборку, которая должна быть репрезентативной - объективно отражать всю генеральную совокупность, т. е. всю популяцию в целом. В обследуемой выборке устанавливают распределение лиц по соответствующим четко очерченным фенотипическим классам, различия между которыми наследственно обусловлены. Затем, исходя из найденных фенотипических частот, определяют генные частоты.
На основе знания генных частот представляется возможность дать описание анализируемой популяции в соответствии с формулой Гарди-Вайнберга и заранее предсказать вероятный характер расщепления в потомстве лиц, относящихся к тем или иным фенотипическим классам. Исследование генных частот имеет важное значение для оценки последствий родственных браков, а также для выяснения генетической истории человеческой популяции в целом.
Частота распространения в популяциях разных аномалий оказывается различной; при этом подавляющее количество соответствующих рецессивных аллелей представлено в гетерозиготном состоянии.

Так, примерно каждый сотый житель Европы гетерозиготен по гену амавротической идиотии (болезнь Шпильмайера-Фогта), тогда как заболевают этой болезнью в юношеском возрасте из 1 млн. только 25 человек, являющихся гомозиготными. Альбиносы в европейских странах встречаются с частотой 1 на 20 000, хотя гетерозиготное состояние этой аллели присуще каждому семидесятому жителю.
Несколько иначе дело обстоит в случае аномалий, наследующихся сцеплено с полом, примером чего может служить дальтонизм - цветная слепота, которая контролируется, по-видимому, рядом аллелей, распределенных по двум тесно сцепленным локусам в Х-хромосоме. Среди мужского населения частота дальтоников (q) соответствует суммарной частоте рецессивных аллелей и составляла, например, в Москве в 30-х годах, по данным Р. И. Серебровской, 7%, в то же время среди женского населения той же популяции цветная слепота была только у 0,5% (q 2), но в гетерозиготном состоянии примерно 13% женщин несут аллели, обусловливающие дальтонизм.
Как мы уже говорили выше, рассматривая генеалогический метод, вероятность появления в потомстве рецессивных гомозигот может быть различной при вступлении в брак лиц, имеющих разную степень родства. Так, у супругов, являющихся по отношению друг к другу двоюродными братьями и сестрами, вероятность рождения детей, гомозиготных по рецессивной аллели, распространенной в популяции с частотой q, составит уже не q 2 , а большую величину, а именно q/16 (1 +15q).
Это связано с тем, что если один из общих предков таких супругов - бабушка или дедушка - нес в гетерозиготе рецессивную аллель, то с вероятностью 1/16 данная аллель передастся обоим двоюродным сибсам.
Вредные последствия родственных браков особенно наглядно проявляются в изолированных популяциях ограниченного размера, так называемых изолятах . Под изолятом понимают группу особей популяции, которые вступают в брак большей частью с особями своей группы и поэтому характеризуются значительным коэффициентом кровного родства. Такими изолятами могут быть отдельные изолированные селения, общины и т. д. Внутри изолята более вероятны родственные браки (инбридинг), и больше шансов на то, что супруги будут нести одинаковые мутантные гены, следствием чего является увеличение вероятности проявления рецессивных аллелей в гомозиготном состоянии. Разные изоляты несут различные концентрации сходных или разных генов.
На Марианских островах и острове Гуам смертность среди местного населения от бокового амиотрофического склероза (связанного с поражением клеток передних рогов спинного мозга) в 100 с лишним раз превышает смертность от этой болезни в других странах. В Южной Панаме в провинции Сан-Блаз весьма заметную часть племени кариба куна составляют альбиносы, которые появляются здесь в каждом поколении. В одном селении на р. Роне в Швейцарии среди 2200 жителей имеется более 50 глухонемых, и еще у 200 обнаруживаются некоторые дефекты слуха. По всей вероятности, во всех подобных случаях резкого увеличения концентрации отдельных аллелей известную роль играет генетический дрифт, неравномерное размножение в прошлом отдельных семей, родов, а также снижение миграции.
По мере роста цивилизации и развития производительных сил общества количество изолятов уменьшается, и их значение для популяции в целом падает. Однако они все еще имеют место.
Знание генных частот, как уже говорилось позволяет предсказывать характер расщепления в потомстве отдельных фенотипических классов родительских особей.
Исходя из формулы Гарди-Вайнберга, можно показать, что при моногенном наследовании расщепление по фенотипу в потомстве доминантных матерей должно осуществляться в соотношении p(1 + pq) доминантов к р рецессивов, или (l+pq):q 2 ; в потомстве рецессивных матерей расщепление по фенотипу должно быть pq 2: q 3 , или p: q.
Приведем пример. В одном исследовании при изучении резус-фактора частота рецессивной аллели rh в популяции составила 0,4, а частота доминантной аллели Rh - 0,6. Отсюда следовало ожидать, что в потомстве резус-положительных матерей частота резус — положительных детей (Rh +) примерно в 7,8 раза будет превышать частоту резус-отрицательных детей (Rh —); в потомстве резус-отрицательных матерей соответствующее превышение будет в 1,5 раза.
Действительные соотношения в обследованной выборке составили:
- в первом случае 1475 Rh + : 182 Rh — , или 8,1: 1,
- во втором случае 204 Rh + : 129 Rh — , или 1,6: 1.
Таким образом, наблюдаемые результаты при расщеплении весьма хорошо соответствуют теоретически ожидаемым результатам, предсказанным на основе анализа генных частот.
Популяционный анализ полиморфизма по группам крови интересен тем, что он помогает понять динамику генетической структуры различных популяций и способствует выявлению связей между ними.
Разные популяции существенно различаются по своей генетической структуре, в частности по группам крови. При этом удается проследить некоторые вполне четкие закономерности. Если концентрация аллели I B наибольшая в районе Индии и Китая, то к востоку и западу от этого района происходит постепенное падение ее вплоть до нуля среди коренных обитателей Америки и Австралии. В то же время у американских индейцев (и аборигенов Австралии и Полинезии) максимума достигает концентрация аллели I 0 . Аллель I А редка у коренного населения Америки, а также в Индии, Аравии, тропической Африке, в Западной Европе.
Для объяснения этих различий в генетической структуре популяций недавно была предложена гипотеза, согласно которой решающим фактором отбора в отношении групп крови системы АВ0 явились эпидемии чумы и оспы. Возбудитель чумы Pasteuvella pest is, обладая свойством антигена 0, оказывается наиболее губительным для людей с группой крови 0, поскольку такие лица не способны вырабатывать достаточное количество антител в случае инфекции. По аналогичной причине вирус оспы наиболее опасен для людей с группой крови А. Там, где свирепствовала чума (Индия, Монголия, Китай, Египет), шла интенсивная элиминация аллели I 0 , а там, где особенно свирепствовала оспа (Америка, Индия, Аравия, тропическая Африка), в первую очередь элиминировалась аллель 1 А. В районах Азии, где чума и оспа были эндемичны, наибольшую частоту получила аллель 1 в.
В главе 5 мы рассмотрели моногенное наследование серповидноклеточной анемии, обусловленное расщеплением по аллелям гена S. Высокая концентрация аллели S в поясе эндемичной малярии (Африка, Средиземноморье) оказалась связанной с повышенной устойчивостью к малярии гетерозигот (Ss) и с возникновением. в результате этого системы сбалансированного наследственного полиморфизма.
Таким образом, в обоих приведенных примерах анализа полиморфизма по группам крови и серповидно-клеточной анемии мы видим, как применение популяционного метода позволяет вскрывать генетическую структуру человеческих популяций.
Онтогенетический метод
Онтогенетический метод позволяет устанавливать по фенотипу носительство рецессивных аллелей в гетерозиготном состоянии и хромосомных перестроек.
Генетической основой проявления рецессивных генов в гетерозиготном состоянии является, по-видимому, неполный блок в цепи синтеза того или иного метаболита, вызванного действием доминантной аллели данного гена.
Известно, что некоторые наследственные болезни проявляются не только у лиц, гомозиготных по аллелям, вызывающим заболевание, но в стертой форме и у гетерозигот. Поэтому в настоящее время усиленно разрабатываются методы определения гетерозиготного носительства в онтогенезе. Так, гетерозиготный носитель фенилкетонурии (повышенное содержание фенилаланина в крови определяется дополнительным введением фенилаланина и последующим определением уровня его (или тирозина) в плазме крови. Наличие гетерозиготности по данной аллели устанавливается по повышенному содержанию фенилаланина. В норме (т. е. у гомозигот по доминантной аллели) уровень фенилаланина не изменяется. В норме в крови присутствует фермент каталаза, необходимый для углеводного обмена, но встречается ген, который в гомозиготном состоянии вызывает отсутствие каталазы. У гомозиготных носителей этого гена наблюдается болезнь акаталаземия - расстройство углеводного обмена. Гетерозиготы занимают промежуточное положение по активности каталазы без большого захождения между доминантными и рецессивными гомозиготами.
По активности каталазы можно точно определить гетерозиготных и гомозиготных носителей аллели акаталаземии среди близких родственников и родителей.
Гетерозиготное носительство аллели, определяющей мышечную дистрофию типа Дюшена, тестируется по активности криатинфосфокиназы. Теперь разработаны, подобные тесты для 40 наследственных болезней, определяемых рецессивными аллелями.
В настоящее время онтогенетический метод обогатился за счет биохимических, иммунологических и молекулярных приемов исследования, описанию которых посвящен ряд специальных руководств.
Важность онтогенетического метода очевидна для установления носительства рецессивного гена в гетерозиготном состоянии у родственников семьи, в которой появляется наследственно больной ребенок. Диагностика в онтогенезе важна для расчета вероятности появления наследственно больных потомков при родственных и смешанных браках. По мере упрощения тестирования гетерозиготного носительства этот метод должен будет внедряться в целях консультации супружеских пар относительно возможности появления заболевания у их детей, а также для изучения распространения мутаций в популяциях.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Генеалогический метод
Генеалогический метод заключается в анализе родословных и позволяет определить тип наследования (доминантный
рецессивный, аутосомный или сцепленный с полом) признака, а также его моногенность или полигенность. На основе полученных сведений прогнозируют вероятность проявления изучаемого признака в потомстве, что имеет большое значение для предупреждения наследственных заболеваний.
При аутосомном наследовании признак характеризуется равной вероятностью проявления у лиц обоих полов. Различают аутосомно-доминантное и аутосомно-рецессивное наследование.
При аутосомно-доминантном наследовании доминантный аллель реализуется в признак как в гомозиготном, так и в гетерозиготном состоянии. При наличии хотя бы у одного родителя доминантного признака последний с разной вероятностью проявляется во всех последующих поколениях. Однако для доминантных мутаций характерна низкая пенетрантность. В ряде случаев это создает определенные трудности для определения типа наследования.
При аутосомно-рецессивном наследовании рецессивный аллель реализуется в признак в гомозиготном состоянии. Рецессивные заболевания у детей встречаются чаще при браках между фенотипически нормальными гетерозиготными родителями. У гетерозиготных родителей (Аа х Аа) вероятность рождения больных детей (аа) составит 25%, такой же процент (25%) буду здоровы (АА), остальные 50% (Аа) будут также здоровы, но окажутся гетерозиготными носителями рецессивного аллеля. В родословной при аутосомно-рецессивном наследовании заболевание может проявляться через одно или несколько поколений.
Интересно отметить, что частота появления рецессивного потомства значительно повышается при близкородственных браках, так как концентрация гетерозиготного носительства у родственников значительно превышает таковую в общей массе населения.
Сцепленное с полом, наследование характеризуется, как правило, неравной частотой встречаемости признака у индивидуумов разного пола и зависит от локализации соответствующего гена в Х- или Y-хромосоме. В X- и Y-хромосомах человека имеются гомологичные участки, содержащие парные гены. Гены, локализованные в гомологичных участках, наследуются так же, как и любые другие гены, расположенные в аутосомах. По-видимому, негомологичные гены имеются и в Y-хромосоме. Они передаются от отца к сыну и проявляются только у мужчин (голандрический тип наследования) .
У человека в Y-хромосоме находится ген, обусловливающий дифференцировку пола. В Х-хромосоме имеется два негомологичных участка, содержащих около 150 генов, которым нет аллельных в Y-хромосоме. Поэтому вероятность проявления рецессивного аллеля у мальчиков более высока, чем у девочек. По генам, локализованным в половых хромосомах, женщина может быть гомозиготной или гетерозиготной. Мужчина, имеющий только одну Х-хромосому, будет гемизиготным по генам, которым нет аллелей в Y-хромосоме.
Наследование, сцепленное с Х-хромосомой, может быть доминантным и рецессивным (чаще рецессивным). Рассмотрим Х - сцепленное рецеcсивное наследование на примере такого заболевания xеловека, как гемофилия (нарушение свертывания крови). Известный всему типу пример: носитель гемофилии королева Виктория была гетерозиготной и передала мутантный ген сыну Леопольду и двум дочерям. Эта болезнь проникла в ряд королевских домов Европы и попала в Россию.
Популяционный метод
Методы генетики популяций широко применяют в исследованиях человека. Внутрисемейный анализ заболеваемости неотделим от изучения наследственной патологии как в отдельных странах, так и в относительно изолированных группах населения. Изучение частоты генов и генотипов в популяциях составляет предмет популяционно-генетического исследования. Это дает информацию о степени гетерозиготности и полиморфизма человеческих популяций, выявляет различия частот аллелей между разными популяциями.
Считают, что закон Харди - Вайнберга свидетельствует о том, что наследование как таковое не меняет частоты аллелей в популяции. Этот закон вполне пригоден для анализа крупных популяций, где идет свободное скрещивание. Сумма частот аллелей одного гена, согласно формуле Харди - Вайнберга р+q=1, в генофонде популяции является величиной постоянной. Сумма частот генотипов аллелей данного гена p2+2pq+q2=1 также величина постоянная. При полном доминировании, установив в данной.популяции число рецессивных гомозигот (q2 - число гомозиготных "особей по рецессивному гену с генотипом аа), достаточно извлечь квадратный корень из полученной величины, и мы найдем частоту рецессивного аллеля а. Частота доминантного аллеля А составит р = 1 - q. Вычислив таким образом частоты аллелей а и А, можно определить частоты соответствующих генотипов в популяции (р2=АА; 2рq=Аа). Например, по данным ряда ученых, частота альбинизма (наследуется как аутосомный рецессивный признак) составляет 1:20 000 (q2). Следовательно, частота аллеля a в генофонде будет q2=l/20000 = /l4l и тогда частота аллеля А будет
p=1-q. p=1. p=1 – 1/141=140/141.
В этом случае частота гетерозиготных носителей гена альбинизма (2pq) составит 2(140/141) x (1/141) = 1/70, или 1,4%
Статистический анализ распространения отдельных наследственных признаков (генов) в популяциях людей в разных странах позволяет определить адаптивную ценность конкретных генотипов. Однажды возникнув, мутации могут передаваться потомству на протяжении многих поколений. Это приводит к полиморфизму (генетической неоднородности) человеческих популяций. Среди населения Земли практически невозможно (за исключением однояйцевых близнецов) найти генетически одинаковых людей. В гетерозиготном состоянии в популяциях находится значительное количество рецессивных аллелей (генетический груз), обусловливающих развитие различных наследственных заболеваний. Частота их возникновения зависит от концентрации рецессивного гена в популяции и значительно повышается при заключении близкородственных браков.
Близнецовый метод
Этот метод используют в генетике человека для выяснения степени наследственной обусловленности исследуемых признаков. Близнецы могут быть однояйцевыми (образуются на ранних стадиях дробления зиготы, когда из двух или реже из большего числа бластомеров развиваются полноценные организмы). Однояйцевые близнецы генетически идентичны. Когда созревают и затем оплодотворяются разными сперматозоидами две или реже большее число яйцеклеток, развиваются разнояйцевые близнецы. Разнояйцевые близнецы сходны между собой не более чем братья и сестры, рожденные в разное время. Частота появления близнецов у людей составляет около 1% (1/3 однояйцевых, 2/3 разнояйцевых); подавляющее большинство близнецов является двойнями.
Так как наследственный материал однояйцевых близнецов одинаков, то различия, которые возникают у них, зависят от влияния среды на экспрессию генов. Сравнение частоты сходства по ряду признаков пар одно- и разнояйцевых близнецов позволяет оценить значение наследственных и средовых факторов в развитии фенотипа человека.
Цитогенетический метод
Цитогенетический метод используют для изучения нормального кариотипа человека, а также при диагностике наследственных заболеваний, связанных с геномными и хромосомными мутациями.
Кроме того, этот метод применяют при исследовании мутагенного действия различных химических веществ, пестицидов, инсектицидов, лекарственных препаратов и др.
В период деления клеток на стадии метафазы хромосомы имеют более четкую структуру и доступны для изучения. Диплоидный набор человека состоит из 46 хромосом:
22 пар аутосом и одной пары половых хромосом (XX - у женщин, XY - у мужчин). Обычно исследуют лейкоциты периферической крови человека, которые помещают в специальную питательную среду, где они делятся. Затем готовят препараты и анализируют число и строение хромосом. Разработка специальных методов окраски значительно упростила распознавание всех хромосом человека, а в совокупности с генеалогическим методом и методами клеточной и генной инженерии дала возможность соотносить гены с конкретными участками хромосом. Комплексное применение этих методов лежит в основе составления карт хромосом человека.
Цитологический контроль необходим для диагностики хромо- сомных болезней, связанных с ансуплоидией и хромосомными мутациями. Наиболее часто встречаются болезнь Дауна(трисомия по 21-й хромосоме), синдром Клайнфелтера (47 XXY), синдром Шершевского - Тернера (45 ХО) и др. Потеря участка одной из гомологичных хромосом 21-й пары приводит к заболеванию крови - хроническому миелолейкозу.
При цитологических исследованиях интерфазных ядер со- матических клеток можно обнаружить так называемое тельце Барри, или половой хроматин. Оказалось, что половой хроматин в норме есть у женщин и отсутствует у мужчин. Он представляет собой результат гетерохроматизации одной из двух Х-хромосом у женщин. Зная эту особенность, можно идентифицировать половую принадлежность и выявлять аномальное количество Х-хромосом.
Выявление многих наследствен- ных заболеваний возможно еще до рождения ребенка. Метод пренатальной диагностики заключается в получении околоплодной жидкости, где находятся клетки плода, и в последующем биохимическом и цитологическом определении возможных наследственных аномалий. Это позволяет поставить диагноз на ранних сроках беременности и принять решение о се продолжении или прерывании.
Биохимический метод
Наследственные заболевания, которые обусловлены генными мутациями, изменяющими структуру или скорость синтеза белков, обычно сопровождаются нарушением углеводного, белкового, липидного и других типов обмена веществ. Наследственные дефекты обмена можно диагностировать посредством определения структуры измененного белка или его количества, выявления дефектных ферментов или обнаружения промежуточных продуктов обмена веществ во внеклеточных жидкостях организма (крови, моче, поте и т.д.). Например, анализ аминокислотных последо- вательностей мутационно измененных белковых цепей гемоглобина позволил выявить несколько наследственных дефектов, лежащих в основе ряда заболеваний, - гемоглобинозов. Так, при сер- повидно-клеточной анемии у человека аномальный гемоглобин вследствие мутации отличается от нормального заменой только одной аминокислоты (глутаминовой кислоты на валин).
К методам, широко используемым при изучении генетики человека относятся: генеалогический, популяционно-статистический, близнецовый, метод дерматоглифики, цитогенетический, биохимический, методы генетики соматических клеток.
Генеалогический метод генетики человека
В основе этого метода лежит составление и анализ родословных. Как метод изучения генетики человека генеалогический метод стали применять с начала ХХ столетия, когда выяснилось, что анализ родословных может заменить собой неприменимый к человеку гибридологический метод. При составлении родословной, исходным является человек, родословную которого изучают – пробанд. При составлении родословных таблиц используют условные обозначения, предложенные Г. Юстом в 1931г. С помощью генеалогического метода может быть установлена наследственная обусловленность изучаемого признака, а также тип его наследования (аутосомно-доминантный, аутосомно-рецессивный, Х-сцепленный, Y-сцепленный). При анализе родословных по нескольким признакам может быть выявлен сцепленный характер их наследования, что используется при составлении хромосомных карт. Этот метод позволяет изучить интенсивность мутационного процесса. Он широко используется в медико-генетическом консультировании для прогнозирования потомства.
Близнецовый метод генетики человека
Этот метод заключается в изучении закономерностей наследования признаков в парах одно- и двуяйцевых близнецов. Он предложен в 1875г. Гальтоном первоначально для оценки роли наследственности и среды в развитии психических свойств человека. Этот метод позволяет выявить наследственный характер признака, оценить эффективность действия на организм некоторых внешних факторов. Суть метода заключается в сравнении проявления признака в разных группах близнецов при учете сходства или различия их генотипов. Монозиготные близнецы, развивающиеся из одной оплодотворенной яйцеклетки, генетически идентичны. Поэтому среди монозиготных близнецов наблюдается высокий процент конкордатных пар, в которых признак развивается у обоих близнецов. Сравнение монозиготных близнецов, воспитывающихся в разных условиях постэмбтионального периода, позволяет выявить признаки, в формировании которых существенная роль принадлежит факторам среды. По этим признакам наблюдается дискордантность – различия. Сохранение сходства несмотря различие условий воспитания, свидетельствует о наследственной обусловленности признака.
Установление соотносительной роли наследственности и среды в развитии различных патологических состояний позволяет врачу правильно оценить ситуацию и проводить профилактические мероприятия при наследственной предрасположенности или осуществлять вспомогательную терапию при его наследственной обусловленности.
Трудности близнецового метода генетики человека связаны:
- с относительно низкой частотой рождения близнецов;
- с идентификацией монозиготности близнецов, что имеет большое значение для получения достоверных выводов.
Несмотря на трудоемкость метода и возможность ошибок при определении их монозиготности, высокая объективность выводов делает его одним из широко применяемых методов генетики человека.
Популяционно- статистический метод генетики человека
С помощью популяционно-статистического метода изучают наследственные признаки в больших группах населения, в одном или нескольких поколениях. Существенным моментом при использовании этого метода является статистическая обработка данных. Этим методом можно рассчитать частоту встречаемости в популяции различных аллелей гена, выяснить распространение в ней различных наследственных признаков, в том числе и заболеваний.
При статистической обработке материала, получаемому при обследовании группы населения по интересующему исследователя признаку, основой для выяснения генетической структуры популяции является закон нентического равновесия Харди-Вайнберга. Он отражает закономерность, в соответствии с которой при определенных условиях соотношения аллелей генов и генотипов в генофонде популяции сохраняется неизменным в ряду поколений этой популяции. На основании этого закона, имея данные о частоте встречаемости в популяции рецессивного фенотипа, обладающего гомозиготным генотипом, можно рассчитать частоту встречаемости указанного аллеля в генофонде данного поколения.
Методы дерматоглифики и пальмоскопии - как методы генетики человека
В 1892г. Ф.Гальтоном в качестве одного из методов исследования человека был предложен метод изучения кожных гребешковых узоров пальцев и ладоней, а также сгибательных ладонных борозд. Он установил, что указанные узоры являются индивидуальной характеристикой человека и не изменяются в течении жизни.
В настоящее время установлена наследственная обусловленность кожных узоров, хотя характер наследования окончательно не выяснен.вероятно, признак наследуется по полигенному типу.
Дерматоглифические исследования важны при идентификации близнецов. Изучение людей с хромосомными заболеваниями выявило у них специфические изменения не только рисунков пальцев и ладоней, но и характера основных сгибательных борозд на коже ладоней. Менее изучены дерматоглифические изменения при генных болезнях.
В основном эти методы генетики человека применяют с целью установления отцовства.
Методы генетики соматических клеток
С помощью этих методов изучают наследственность и изменчивость соматических клеток, что компенсирует невозможность применения к человеку гибридологического анализа. Эти методы, основанные на размножении этих клеток в искусственных условиях, анализировать генетические процессы в отдельных клетках организма, и благодаря полноценности генетического материала использовать их для изучения генетических закономерностей целого организма.
В генетических исследованиях человека используют следующие приемы:
- культивирование – позволяет получить достаточное количество генетического материала для различных исследований;
- клонирование – получение потомков одной клетки;
- селекция соматических клеток с помощью искусственных сред используется для отбора клеток с интересующими исследователя свойствами;
- гибридизация соматических клеток основана на слиянии совместно культивируемых клеток разных типов.
Гибридные клетки, содержащие 2 полных генома, при делении обычно «теряют» хромосомы предпочтительно одного из видов. Таким образом, можно получать клетки с желаемым набором хромосом, что дает возможность изучать сцепление генов и их локализацию в определенных хромосомах.
Благодаря методам генетики соматических клеток можно изучать механизмы первичного действия и взаимодействия генов, регуляцию генной активности. Развитие этих методов определило возможность точной диагностики наследственных болезней в пренатальном периоде.
Цитогенетический метод генетики человека
Цитогенетический метод генетики человека основан на микроскопическом изучении хромосом в клетках человека. Его стали широко применять с 1956г. Современный этап в применении цитогенетического метода связан с разработанным в 1969г. Т. Касперсоном методом дифференциального окрашивания хромосом, который расширил возможности цитогенетического анализа. Применение цитогенетического метода позволяет изучать нормальную морфологию хромосом и кариотипа в целом, определять генетический пол организма и диагностировать различные хромосомные болезни, связанные с изменением числа хромосом или с нарушением их структуры.
Материалом для цитогенетических исследований служат клетки человека получаемые из разных тканей. Непременным требованием для изучения хромосом является наличие делящихся клеток (в основном лимфоциты периферической крови). В качестве экспресс-метода, выявляющего изменение числа половых хромосом, используют метод определения полового хроматина в неделящихся клетка слизистой оболочки щеки.
Биохимический метод генетики человека
С помощью биохимических методов изучают наследственные заболевания, обусловленные генными мутациями, и полиформизм по нормальным первичным продуктам генов. Впервые эти методы генетики человека стали применять в начале ХХ в. В последнее время их широко используют в поиске новых форм мутантных аллелей. С их помощью описано более 1000 врожденных болезней обмена веществ. Для многих из них выявлен дефект первичного генного продукта.
Биохимическую диагностику наследственных нарушений обмена проводят в 2 этапа. На первом этапе отбирают предположительные случаи заболеваний, на втором – более сложными и точными методами уточняют диагноз заболевания. Применение биохимических исследований для диагностики заболеваний в пренатальном периоде или непосредственно после рождения позволяет своевременно выявить патологию и начать специфические медицинские мероприятия.
Генетика человека – это наука, которая изучает кроме наследственности и изменчивости формирование нормальных признаков человека.
Предметом являются нормальные признаки человека.
Задачи генетики :
Изучение закономерностей генетической детерминации признаков человека
Изучение материальной структуры наследования генов
Исследование организации потока информации в клетках человека
Анализ характера взаимодействия между генами в процессе формирования признаков
Изучение влияния факторов внешней среды на наследственность человека
История развития генетики человека.
1815г Адамс «Филосовский тракт о наследственных свойствах человеческой расы». 1 справочник о генетики.
1866г Флоренский «Усовершенствование и вырождение человеческого рода»
Влияние внешней среды
Влияние близкородственных браков
Гальтон – основоположник методов генетики: генеалогический, близнецовый, статистический.
Гарред. Алкаптонурия – вырожденная ошибка метаболизма, рецессивное заболевание (биохимическая генетика – начало развития).
Ю.А. Гиритиенко – руководитель первой кафедры генетики в Петрограде в 1919 году.
1932 – 37гг . в Москве открыт первый медико-генетический институт под руководствомЛевиха.
Большую роль сыграли ученые Харди и Вайнберг =>основные положения стабильности популяции.
Особенности генетического анализа у человека.
Бисоциальная природа человека. На формирование любого признака влияют средовые факторы и социальная среда, в которой обитает человек.
Невозможность постановки прямых экспериментов
Невозможность использования гибридологического метода
Позднее наступление половой зрелости => большая продолжительность жизни одного поколения
Малое число потомков
Большое число хромосом (46)
Невозможность создания одинаковых и строго контролируемых условий жизни для потомков
Сущность генеалогического метода и задачи, решаемые с помощью него в генетике человека.
Генеалогический метод – метод составления родословной. В медицине – клинико-генеалогический.
Основан на составлении родословной человека и изучении характера наследования признака. Впервые этот метод был предложен Ф. Гальтоном в 1865 г. Это самый давний метод. Суть его состоит в установлении родословных связей и определении доминантных и рецессивных признаков и характера их наследования. Особенно эффективен этот метод при исследовании генных мутаций.
Установление наследственного характера признака
Определение типа наследования признака
Анализ сцепления генов и картирования хромосом
Изучение интенсивности мутационного процесса
Расшифровка механизмов взаимодействия генов
Использование данного метода при медико-генетическом консультировании
Принципы построения родословных и используемая при этом символика.
Пробанд – человек, обратившийся к врачу-генетику за консультацией
Сибсы – родные братья и сестры пробанда
Использование этого метода возможно в том случае, когда известны прямые родственники - предки обладателя наследственного признака (пробанда ) по материнской и отцовской линиям в ряду поколений или потомки пробанда также в нескольких поколениях. При составлении родословных в генетике используется определенная система обозначений. После составления родословной проводится ее анализ с целью установления характера наследования изучаемого признака.
Условные обозначения, принятые при составлении родословных: 1 - мужчина; 2 - женщина; 3 - пол не выяснен; 4 - обладатель изучаемого признака; 5 - гетерозиготный носитель изучаемого рецессивного гена; 6 - брак; 7 - брак мужчины с двумя женщинами; 8 - родственный брак; 9 - родители, дети и порядок их рождения; 10 - дизиготные близнецы; 11 - монозиготные близнецы.
Благодаря генеалогическому методу были определены типы наследования многих признаков у человека. Так, по аутосомно-доминантному типу наследуются полидактилия (увеличенное количество пальцев), возможность свертывать язык в трубочку, брахидактилия (короткопалость, обусловленная отсутствием двух фаланг на пальцах), веснушки, раннее облысение, сросшиеся пальцы, заячья губа, волчья пасть, катаракта глаз, хрупкость костей и многие другие. Альбинизм, рыжие волосы, подверженность полиомиелиту, сахарный диабет, врожденная глухота и другие признаки наследуются как аутосомно-рецессивные.
Близнецовый метод, его сущность и задачи, решаемые с помощью него в генетике человека.
Близнецовый метод основан на изучении фенотипа и генотипа близнецов для определения степени влияния среды на развитие различных признаков. Этот метод в 1876 г. предложил английский исследователь Ф. Гальтон для разграничения влияния наследственности и среды на развитие различных признаков у человека. Близнецовый метод позволяет определить степень проявления признака у пары, влияние наследственности и среды на развитие признаков. Все различия, которые проявляются у однояйцевых близнецов, имеющих одинаковый генотип, связаны с влиянием внешних условий. Большой интерес представляют случаи, когда такая пара была по каким-то причинам разлучена в детстве и близнецы росли и воспитывались в разных условиях. Изучение разнояйцевых близнецов позволяет проанализировать развитие разных генотипов в одинаковых условиях среды. Близнецовый метод позволил установить, что для многих заболеваний значительную роль играют условия среды, при которых происходит формирование фенотипа. Например, такие признаки как группа крови, цвет глаз и волос определяются только генотипом и от среды не зависят. Некоторые заболевания, хотя и вызываются вирусами и бактериями, в некоторой степени зависят от наследственной предрасположенности. Такие заболевания, как гипертония и ревматизм, в значительной степени определяются внешними факторами и в меньшей степени – наследственностью. Таким образом, близнецовый метод позволяет выявить роль генотипа и факторов среды в формировании признака, для чего изучаются и сравниваются степени сходства (конкордантность) и различий (дискордантность) монозиготных и дизиготных близнецов.
Типы близнецов и их характеристика. Причины и частота рождения близнецов.
Среди близнецов выделяются однояйцевые и двуяйцевые. Однояйцевые близнецы (идентичные) образуются из одной зиготы (монозиготные), разделившейся на ранней стадии дробления на две части. В этом случае одна оплодотворенная яйцеклетка дает начало не одному, а сразу двум зародышам. Они имеют одинаковый генетический материал, всегда одного пола, и наиболее интересны для изучения. Сходство у таких близнецов почти абсолютное. Мелкие различия могут объясняться влиянием условий развития. Двуяйцевые близнецы (неидентичные, или дизиготные) образуются из различных зигот, в результате оплодотворения двух яйцеклеток двумя сперматозоидами. Они похожи друг на друга не более чем родные братья или сестры, рожденные в разное время. Такие близнецы могут быть однополыми и разнополыми.
Общая частота рождения близнецов составляет в среднем 1,1 – 1,2 % от всех рождений; из них около 1/3 приходится на монозиготных близнецов, а 2/3 – на дизиготных близнецов. Частота рождения монозиготных близнецов сходна в разных популяциях, а частота рождения дизиготных близнецов существенно различается в разных популяциях. Например, в США дизиготные близнецы среди негров рождаются чаще, чем среди белых. В Европе дизиготные близнецы рождаются с частотой 8 на 1 000 рождений. Самая низкая частота рождения дизиготных близнецов присуща монголоидным популяциям, где она составляет 2-2,5 на 1 000 рождений. Вероятность рождения дизиготных близнецов повышается с увеличением возраста матери, а также порядкового номера рождения детей. Это правило касается исключительно дизиготных близнецов. Влияние возраста матери объясняется, по-видимому, повышением с возрастом уровня фолликулостимулирующего гормона у женщин. Фолликулостимулирующие гормон – гормон передней доли гипофиза, который стимулирует образование фолликулов в яичниках, их рост и созревание, способствует процессу выбора доминантного фолликула и образованию зрелыхграафовых пузырьков. Повышение уровня этого гормона и приводит к более частой полиовуляции. Эту гипотезу подтверждают и факты повышенной частоты многоплодных родов у женщин, проходивших лечение от бесплодия с помощью гонадотропных гормонов. В отношении дизиготного многоплодия имеются также факты, свидетельствующие о влиянии генетических факторов на вероятность рождения близнецов.
Вероятность рождения дизиготных близнецов выше для тех женщин, родственники которых уже имели близнецов. Возможно, основной генетически детерминированной причиной в этом случае также может быть уровень гонадотропина. В отношении монозиготных близнецов таких данных не имеется. В связи с несколько повышенной смертностью среди близнецов, по сравнению с таковой у одиночно рожденных, доля близнецов среди населения составляет всего 0,9 %. Столь низкая частота близнецов осложняет подбор достаточного количества пар с исследуемым признаком.
Оценка роли генетических и средовых факторов в формировании качественных и количественных признаков по данным близнецовых исследований.
Для оценки эффективности воздействия некоторых внешних факторов Н + Е=1 Н = Сmz – Cdz/100 – Cdz где Н- наследственный фактор С – средовой фактор Сmz – монозиготы Сdz – дизиготы Если Н = ближе к 0 –большее влияние оказывают средовые факторы Если Н = 1 – 0,7 – генетические Если Н = 0,4 – 0,7 – больше средовые, чем генетические
Сущность популяционно-статистического метода и задачи, решаемые с помощью него в генетике человека.
Это метод изучения распространения наследственных признаков (наследственных заболеваний) в популяциях. Существенным моментом при использовании этого метода является статистическая обработка получаемых данных. Под популяцией понимают совокупность особей одного вида, длительное время обитающих на определенной территории, свободно скрещивающихся друг с другом, имеющих общее происхождение, определенную генетическую структуру и в той или иной степени изолированных от других таких совокупностей особей данного вида. Популяция является не только формой существования вида, но и единицей эволюции, поскольку в основе микроэволюционных процессов, завершающихся образованием вида, лежат генетические преобразования в популяциях.
Изучением генетической структуры популяций занимается особый раздел генетики - популяционная генетика.
Одним из важных направлений в современной генетике является популяционная генетика. Она изучает генетическую структуру популяций, их генофонд, взаимодействие факторов, обусловливающих постоянство и изменение генетической структуры популяций. Под популяцией в генетике понимается совокупность свободно скрещивающихся особей одного вида, занимающих определенный ареал и обладающих общим генофондом в ряду поколений. (Генофонд - это вся совокупность генов, встречающихся у особей данной популяции).
В медицинской генетике популяционно-статистический метод используется при изучении наследственных болезней населения, частоты нормальных и патологических генов, генотипов и фенотипов в популяциях различных местностей, стран и городов. Кроме того, этот метод изучает закономерности распространения наследственных болезней в разных по строению популяциях и возможность прогнозировать их частоту в последующих поколениях.
Популяционно-статистический метод используется для изучения:
а) частоты генов в популяции, включая частоту наследственных болезней;
б) закономерности мутационного процесса;
Закон Хайди-Вайнберга. Условия идеальной популяции.
Для выяснения частот встречаемости тех или иных генов и генотипов используется закон Харди-Вайнберга .
Закон Харди-Вайнберга
В идеальной популяции из поколения в поколение сохраняется строго определенное соотношение частот доминантных и рецессивных генов (1), а также соотношение частот генотипических классов особей (2).
p + q = 1, (1) р 2 + 2pq + q 2 = 1, (2)
где p - частота встречаемости доминантного гена А ; q - частота встречаемости рецессивного гена а ; р 2 - частота встречаемости гомозигот по доминанте АА ; 2pq - частота встречаемости гетерозигот Аа ; q 2 - частота встречаемости гомозигот по рецессиву аа .
Идеальной популяцией является достаточно большая, панмиктическая (панмиксия - свободное скрещивание) популяция, в которой отсутствуют мутационный процесс, естественный отбор и другие факторы, нарушающие равновесие генов. Понятно, что идеальных популяций в природе не существует, в реальных популяциях закон Харди-Вайнберга используется с поправками.
Закон Харди-Вайнберга, в частности, используется для примерного подсчета носителей рецессивных генов наследственных заболеваний. Например, известно, что в данной популяции фенилкетонурия встречается с частотой 1:10000. Фенилкетонурия наследуется по аутосомно-рецессивному типу, следовательно, больные фенилкетонурией имеют генотип аа , то есть q 2 = 0,0001. Отсюда: q = 0,01; p = 1 - 0,01 = 0,99. Носители рецессивного гена имеют генотип Аа , то есть являются гетерозиготами. Частота встречаемости гетерозигот (2pq ) составляет 2 · 0,99 · 0,01 ≈ 0,02. Вывод: в данной популяции около 2% населения - носители гена фенилкетонурии. Заодно можно подсчитать частоту встречаемости гомозигот по доминанте (АА ): p 2 = 0,992, чуть меньше 98%.
Изменение равновесия генотипов и аллелей в панмиктической популяции происходит под влиянием постоянно действующих факторов, к которым относятся: мутационный процесс, популяционные волны, изоляция, естественный отбор, дрейф генов, эмиграция, иммиграция, инбридинг. Именно благодаря этим явлениям возникает элементарное эволюционное явление - изменение генетического состава популяции, являющееся начальным этапом процесса видообразования.
Приведите формулы для расчета частот генов и генотипов по эритроцитарным изоантигенам в популяции человека (системы АВ0, резус, MN ).
Рассмотрим случай, когда один ген 0 рецессивен по отношению к двум другим - А и В , которые кодоминантны по отношению друг к другу. В конкретной популяции наблюдаем следующие соотношения:
|
Фенотип |
Генотип |
Частота генотипа |
|
r 2 |
||
|
p 2 + 2pr |
||
|
q 2 + 2pr |
||
Оценка частот (p, q, r ) генов А , В , 0 проводится по известным формулам Ф. Бернштейна следующим образом.
Находим предварительные оценки частот генов 0 , А и В :
r" = (0" ) 1/2 ;
p" = 1 - (A" + 0" ) 1/2 ; (6 )
q" = 1 - (B" + 0" ) 1/2 ;
где 0" , A" и B" - частоты фенотипов, т.е. отношение числа лиц с определенным фенотипом к объему выборки.
Когда сумма предварительных частот генов не равна 1, вводим поправку D = 1 - (r" + p" + q" ).
r = (r" + 1/2D ) (1 + 1/2D );
p = p" (1 + 1/2D );
q = q" (1 + 1/2D ); (7 )
r + p + q = 1.
Эти формулы справедливы для оценки частот генов групп крови АВ0, а также системы вкусовой чувствительности к РТС при выявлении в группе обследованных гиперсенситивных лиц.
Биохимический метод и его использование в генетике человека.
Позволяет обнаружить нарушения в обмене веществ, вызванные изменением генов и, как следствие, изменением активности различных ферментов. Наследственные болезни обмена веществ подразделяются на болезни углеводного обмена (сахарный диабет), обмена аминокислот, липидов, минералов и др.
Фенилкетонурия относится к болезням аминокислотного обмена. Блокируется превращение незаменимой аминокислоты фенилаланин в тирозин, при этом фенилаланин превращается в фенилпировиноградную кислоту, которая выводится с мочой. Заболевание приводит к быстрому развитию слабоумия у детей. Ранняя диагностика и диета позволяют приостановить развитие заболевания.
Причиной многих врожденных нарушений метаболизма являются различные дефекты ферментов, возникающие вследствие изменяющих их структуру мутаций. Биохимичские показатели (первичный продукт гена, накопление патологических метаболитов внутри клетки и во всех клеточных жидкостях больного) более точно отражают сущность болезни по сравнению с показателями клиническими, поэтому их значение в диагностике наследственных болезней постоянно возрастает. Использование современных биохимических методов (электрофореза, хроматографии, спектроскопии и др.) позволяют определять любые метаболиты, специфические для конкретной наследственной болезни.
Предметом современной биохимической диагностики являются специфические метаболиты, энзимопатии, различные белки.
Объектами биохимического анализа могут служить моча, пот, плазма и сыворотка крови, форменные элементы крови, культуры клеток (фибробласты, лимфоциты).
Для биохимической диагностики используются как простые качественные реакции (например, хлорид железа для выявления фенилкетонурии или динитрофенилгидразин для выявления кетокислот), так и более точные методы.
Сущность цитогенетического метода и его применение в генетике человека.
Основу метода составляет микрооскопическое изучение кариотипа. Кариотип – это совокупность признаков хромосомного набора соматической клетки организма (форма хромосом, их количество, размеры).
Цитогенетический метод заключается в микроскопическом исследовании структуры хромосом и их количества у здоровых и больных людей. Из трех типов мутаций под микроскопом могут обнаруживаться лишь хромосомные и геномные мутации. Наиболее простым методом является экспресс-диагностика – исследование количества половых хромосом по Х-хроматину. В норме у женщин одна Х-хромосома в клетках находится в виде тельца хроматина, а у мужчин такое тельце отсутствует. При трисомии по половой паре у женщин наблюдаются два тельца, а у мужчин – одно. Для идентификации трисомии по другим парам исследуется кариотип соматических клеток и составляется идиограмма, которая сравнивается со стандартной.
В 1959 г. французские ученые Д. Лежен, Р.Тюрпен и М. Готье установили хромосомную природу болезни Дауна. В последующие годы были описаны многие другие хромосомные синдромы, часто встречающиеся у человека. Цитогенетика стала важнейшим разделом практической медицины. В настоящее время цитогенетический метод применяется для диагностики хромосомных болезней, составления генетических карт хромосом, изучения мутационного процесса и других проблем генетики человека.
В 1960 г. в г. Денвере (США) была разработана первая Международная классификация хромосом человека. В ее основу легли размеры хромосом и положение первичной перетяжки - центромеры.
Дерматоглифический метод и его использование в генетике человека.
Он основан наизучении кожных гребешковых узоров пальцев и ладоней, а также сгибательных ладонных борозд (2, 11). Характер наследования гребневого счета (число линий в узоре на отдельных пальцах) и папиллярные узоры определяются генотипом, что позволяет диагностировать на ранних этапах онтогенеза ряд патологий и определять их природу. Впервые дерматоглифический метод в генетике был предложен в 1892 году Ф.Гальтоном. Именно он установил, что указанные узоры не меняются в течение жизни и являются индивидуальной характеристикой человека. Гальтон уточнил и дополнил классификацию рельефа кожных узоров, основы которой разработаны были Пуркинье еще в 1823 году. Позднее классификацию Ф. Гальтона усовершенствовали, и сейчас она широко применяется в криминалистике и генетических исследованиях. В 1939г. впервые описаны дерматоглифы при синдроме Дауна. Это исследование положило начало описанию дерматоглифов при других хромосомных болезнях: синдромах Клайнфельтера, Шерешевского- Тернера, синдроме «кошачьего крика», что позволило использовать методы дерматоглифики и пальмоскопии в диагностике этих заболеваний. Описаны специфические отклонения этих показателей при шизофрении, миастении, лимфоидной лейкемии. Таким образом, современная наука обладает большим арсеналом методов, позволяющих получить полные знания о наследственности человека и выявить наследственную изменчивость. Однако ряд изменений человеческих признаков носят ненаследственную природу и являются модификациями . Они отражают изменение фенотипа под действием средовых факторов. Возможность организма варьировать степень изменчивости признаков носит название нормы реакции . Норма реакции организма определяется генотипом и может быть широкой или узкой. Люди различаются, например, нормой реакции на инсоляцию – облучение солнечными лучами: кожа одних людей приобретает темный загар, а кожа других при той же дозе облучения – обгарает. У жителей высокогорий уровень гемоглобина на 30% выше, чем у жителей долин. При подъеме в горы у людей увеличивается содержание гемоглобина как адаптивное приспособление к пониженному содержанию кислорода, при возвращении в долину содержание гемоглобина вновь снижается. Знание нормы реакции организма позволяет подбирать оптимальные условия для проявления тех или иных признаков и управлять изменчивостью.
Молекулярно-генетический метод генетики человека.
Они связаны с выделением молекул ДНК из отдельных хромосом, либо митохондрий, с последующим изучением структуры этих молекул, выявлении изменений в определенных участках гена. Это позволяет проводить молекулярную диагностику наследственной патологии. Полученные этими методами данные позволяют получить более полные представления о геноме человека
Конечный итог молекулярно-генетических методов - выявление изменений в определенных участках ДНК, гена или хромосомы. В их основе лежат современные методики работы с ДНК или РНК. В 70-80 гг. в связи с прогрессом в молекулярной генетике и успехами в изучении генома человека молекулярно-генетический подход нашел широкое применение.
Начальным этапом молекулярно-генетического анализа является получение образцов ДНК или РНК. Для этого используют геномную ДНК (вся
ДНК клетки) или отдельные ее фрагменты. В последнем случае, чтобы получить достаточное количество таких фрагментов, необходимо, амплифицировать (размножить) их. Для этого пользуются полимеразной цепной реакцией - быстрым методом ферментативной репликации определенного фрагмента ДНК. С его помощью можно амплифицировать любой участок ДНК, расположенный между двумя известными последовательностями.
Анализировать огромные молекулы ДНК в том виде, в котором они существуют в клетке, невозможно. Поэтому прежде их необходимо разделить на части, обработать разнообразными рестриктазами - бактериальными эндонуклеазами. Эти ферменты способны разрезать двойную спираль ДНК, причем места разрыва строго специфичны для данного образца.
Методы генетики соматических клеток.
Тот факт, что соматические клетки несут в себе весь объем генетической информации, дает возможность изучать на них генетические закономерности всего организма.
Основу метода составляет культивирование отдельных соматических клеток человека и получение из них клонов, а так же их гибридизацию и селекцию.
Соматические клетки обладают рядом особенностей:
Быстро размножаются на питательных средах;
Легко клонируются и дают генетически однородное потомство;
Клоны могут сливаться и давать гибридное потомство;
Легко подвергаются селекции на специальных питательных средах;
Клетки человека хорошо и долго сохраняются при замораживании.
Соматические клетки человека получают из разных органов - кожи, костного мозга, крови, ткани эмбрионов. Однако чаще всего используют клетки соединительной ткани (фибробласты) и лимфоциты крови.
С помощью метода гибридизации соматических клеток:
а) изучают метаболические процессы в клетке;
б) выявляют локализацию генов в хромосомах;
в) исследуют генные мутации;
г) изучают мутагенную и канцерогенную активность химических веществ.
С помощью этих методов изучают наследственность и изменчивость соматических клеток, что в значительной мере компенсирует невозможность применения к человеку метода гибридологического анализа.
Методы генетики соматических клеток, основанные на размножении этих клеток в искусственных условиях, позволяют не только анализировать генетические процессы в отдельных клетках организма, но благодаря полноценности наследственного материала, заключенного в них, использовать их для изучения генетических закономерностей целостного организма.
В связи с разработкой в 60-х гг. XX в. методов генетики соматических клеток человек оказался включенным в группу объектов экспериментальной генетики. Благодаря быстрому размножению на питательных средах соматические клетки могут быть получены в количествах, необходимых для анализа. Они успешно клонируются, давая генетически идентичное потомство. Разные клетки могут, сливаясь, образовывать гибридные клоны. Они легко подвергаются селекции на специальных питательных средах и долго сохраняются при глубоком замораживании. Все это позволяет использовать культуры соматических клеток, полученные из материала биопсий (периферическая кровь, кожа, опухолевая ткань, ткань эмбрионов, клетки из околоплодной жидкости), для генетических исследований человека, в которых используют следующие приемы: 1) простое культивирование, 2) клонирование, 3) селекцию, 4) гибридизацию.











